珍珠黄杨,拉丁名 Buxus Sinica var.Parvifolia. 别名小老树,鱼鳞木。黄杨科,黄杨属,原仅间断分布于一些中国的亚热带高山区域,生长缓慢,自然形态优美,属我国特有的世界濒危珍贵树种。根据实地考察,珍珠黄杨正面临着自然环境和人类活动所带来的双重压力,特别是过去几十年中的非法采挖和生境破坏,致使其种群锐减。由于生境不断地片断化和种群间交流的日益困难,珍珠黄杨局限在一些狭小的孤立区域内,最终被分割成岛屿状小亚群体。生境片断化的研究显示,这种过程最终不仅将导致群体的消退和遗传变异的丧失,而且由此导致的适宜生境的最小化,增加了遗传上关系紧密个体间的交配机会。珍珠黄杨是个自交可亲和物种,风媒为主,兼有虫媒。而自交的后果就是遗传漂变。珍珠黄杨生境之间距离的扩大更增加了基因交流难度,近交加剧,遗传重组减少,遗传多样性降低,自然繁衍困难。目前,文献记载的珍珠黄杨许多分布点已经找不到天然群体,显然部分遗传资源已经丢失。不仅如此,一些分布点的盗挖甚至公开交易仍在持续,由于野生珍珠黄杨原生地生境特殊,下山后罕见存活。如果不能采取有效措施来遏制其人类残害、生态环境退化以及恶劣自然灾害的侵袭,野生资源最终有可能灭绝。

关键词:表型特征 野生资源 濒危 生长特性 保护利用 引种 驯化 继代扦插 选择育种

珍珠黄杨,拉丁名 Buxus Sinica var Parvifolia. 别名小老树,鱼鳞木。黄杨科,黄杨属。为中国特有的濒危珍稀观赏树种,原仅零星分布于中国华东的一些亚热带高山区域。自20世纪后期,在其他地方先后发现少些与之叶片相近的野生品种,和在低海拔地区以种子繁殖的栽培品种,虽然都泛称为珍珠黄杨,但与珍珠黄杨华东特有种相比有着很大的差异。多年的试验及调研结果表明,由于珍珠黄杨遗传多样性丰富复杂,对环境较为敏感,通过种子繁殖的后代极易产生叶片变大,植株变高,枝条变长,树型松散等"逆向变异",从而失去其本有的形态特征和应有的观赏价值。而通过无性繁殖,虽然保留了珍珠黄杨的观赏特性,生长特性却难以改变。并且,除新近培育成功的“好雨”新品种以外,其他几乎所有珍珠黄杨类型,对环境要求均较高,尤其是生长过程中枝条萎缩现象,也很难改变。
总的来说,珍珠黄杨华东特有种具有以下主要特征:生长缓慢、生长态势较弱,有主干或不明显,株型成开心形,植株低矮。观赏性较高,但适应性较弱。表形性状为:常绿灌木或小乔木,树皮淡褐或灰白色,木质细腻坚硬,分枝短而密集,小枝具四菱形,节间长5 -7mm。叶对生,叶片先端圆或稍尖,倒卵形或椭圆形,长不及1 cm,宽6-8 mm,全缘,革质有光泽,幼叶期为翠绿色,成熟后深绿色,深秋及冬季呈深褐色,基部宽楔形,侧脉明显隆起。花序成头状簇生于叶腋或枝端,轴长1-2mm,序期6月起,第二年4月开放,花淡黄色,苞片阔卵形. 长约2mm,外萼片卵状椭圆形,内萼片近圆形,长2mm,子房较花柱稍长,花柱粗扁,柱头倒心形,下延达花柱中部。蒴果近球形,紫褐色,直径6-7 mm,果期7月。
有学者认为,珍珠黄杨是瓜子黄杨的变种。但好雨园景多年的研究试验表明,其在低海拔环境下以种子繁殖的后代,表形特征几乎均与普通小叶黄杨无异,因此对珍珠黄杨起源的结论尚需做进一步的研究。
好雨珍珠黄杨,简称好雨黄杨。是江苏如皋好雨园景翟锦如先生,在通过对华东野生珍珠黄杨特有种进行引种、驯化和培育的基础上,于2016年选育而成的珍珠黄杨无性系新品种。因具有珍珠黄杨其类型无可替代的诸多优良特性,且最初发现于如皋好雨园景,故而得名。好雨珍珠黄杨的主要特点,是在保留了其他珍珠黄杨无性系所具有叶片小、枝节短、自然成型等优良形态特征的前提下,摒弃了珍珠黄杨生长过于缓慢、适应性弱、容易枯枝萎缩等生长缺陷,并且分蘖性强、易成型,抗逆性强、养护简便,耐阴性强、可室内放置。株型矮小茂盛,叶色四季变化鲜明,自然形态更为优美。好雨珍珠黄杨,不仅是制作盆景、点缀山石、建造袖珍园林的极好材料。同时,其叶片密集常绿,平展耐荫,能有效吸收电脑等有害紫外线辐射;株型小中见大,近处见远,能调节瞳孔,消除眼疲劳,从而有效减缓眼近视的发生。好雨珍珠黄杨的培育成功,必将在被人类广泛利用的同时,从根本上改变珍珠黄杨的濒危状态。

黄杨科属于泛热带成分,起源于古南大陆,分布遍及东西两半球的热带地区及亚热带甚至温带。黄杨科全世界共有4属,约100种(郑勉等,1980)。我国已知约17种及几个亚种和变种,西自西藏,东至台湾,南达海南岛,西北至甘肃南部均产,但主要分布于我国西部及西南部。
珍珠黄杨作为一种狭域性物种,原仅间断分布于一些中国的亚热带高山区域。江西三清山1920m,庐山五老峰七星衡,福建崇安黄岗山,浙江龙塘山海拔1600 m、昌化海拔1700 m,安徽大别山区潜山水吼岭海拔1400-1700 m,天柱峰海拔1200-1600 m、舒城晓天猪头尖海拔1200 m、歙县清凉峰海拔1200-1720 m、黄山海拔600-1600 m。珍珠黄杨多生于岩石缝隙间,一般东坡纯林分布,而西坡混生。
根据国际自然与资源保护联盟的濒危物种等级新标准,珍珠黄杨属濒危种。理由是:1.现有分布区面积小,仅在一些高山地区零星分布。2.地理分布面积有持续减少迹象。如清凉峰三个自然保护区目前仅存一个珍珠黄杨的分布点,文献报道江西庐山、安徽万佛山有分布,但现在在当地野外已基本灭绝。并且从已调查的一些野外分布群来看,除少数生境外,均处于种群密度小,幼龄个体数量少,种群年龄结构老化,缺乏后续资源的衰退状态。整个分布区有向高海拔地区退缩的趋势。虽然,过去有极少高山地区新发现珍珠黄杨的报道。无论怎样,珍珠黄杨分布区极为狭小,种群数量在持续减少是可以肯定的。
种质资源保存的主要依据是种内遗传多样性,物种遗传多样性是其生存及进化的前提和基础。遗传多样性越丰富,在环境发生变化时生存下来的可能性就越大,遗传多样性丧失将对其生存带来不利的影响。珍珠黄杨尽管目前检测出较高的遗传多样性,但其多样性是建立在对各种微生境十分依赖,特化程度很高的基础上,一旦其生境发生剧烈变化,很难重新适应变化后的环境,从而发生灭绝。
根据实地考察,珍珠黄杨正面临着自然环境和人类活动所带来的双重压力,特别是过去几十年中的非法采挖和生境破坏, 致使其种群锐减。由于生境不断地片断化和种群间交流的日益困难,珍珠黄杨局限在一些狭小的孤立区域内,最终被分割成岛屿状小亚群体。生境片断化的研究显示,这种过程最终不仅将导致群体的消退和遗传变异的丧失,而且由此导致的适宜生境的最小化,增加了遗传上关系紧密个体间的交配机会。珍珠黄杨是个自交可亲和物种,风媒为主,兼有虫媒。而自交的后果就是遗传漂变。珍珠黄杨生境之间距离的扩大更增加了基因交流难度,近交加剧,遗传重组减少,遗传多样性降低,自然繁衍困难。目前,文献记载的珍珠黄杨许多分布点已经找不到天然群体,显然部分遗传资源已经丢失。不仅如此,一些分布点的盗挖甚至公开交易仍在持续,由于野生珍珠黄杨原生地生境特殊,下山后罕见存活。如果不能采取有效措施来遏制其人类残害、生态环境退化以及恶劣自然灾害的侵袭,野生资源最终有可能灭绝。

中国是世界上具有生物多样性的国家之一,丰富的植物资源,特别是众多的中国特有的资源植物是我们无比宝贵的财富。然而,中国也是生物多样性受威胁最严重的国家之一,自然灭绝、过度利用及生境破坏使得许多植物失去“家园”,稀有濒危植物正以每天1种的速度走向灭绝。全球现约有30万种植物,中国占其中的三分之一。但中国的3万多种高等植物中约有20%正处于稀有濒危状态。目前中科院在全国有12个植物园,各地还有140多个植物园,但这些植物园仅保护了不到一半的中国植物资源。
保护珍稀濒危植物具有多方面的价值:珍稀濒危植物是研究植物起源、系统进化的有利证据,是植物遗传育种的珍贵材料,是植物生殖生态研究的可靠借鉴,是对未知种研究的直接参照物,是古地质、古气候的生动记录,并且具有很高的经济价值和文化价值。
植物野生型是物种的主要基因库,栽培型是野生型人工定向选择的结果。一般情况下,在选择过程中遗传多样性丧失是不可避免的,野生型比栽培型具有高的遗传多样性,研究结果证实了这一点。但是好雨珍珠黄杨种质资源栽培群体的遗传多样性与天然群体比较接近,究其原因可能是在引种驯化过程中部分个体产生了适应性进化,产生了突变,再通过人工选择隔离,使得变异保存了,所以在生长环境趋于均质的情况下仍维持了较高的遗传多样性。但是如果不能有效地保存稀有栽培类型,栽培群体的遗传资源可能存在丢失的危险。 未来如果能在有效控制成本的前提下扩大人工繁殖规模,将不仅能保证其异地保护和可持续存活,也会促进其原地保护甚至在适宜的自然环境中进行回归引种...
人类的利用从某种程度上说是最好的保护。
众所周知,世界的盆景艺术发源于我国,隋唐时期传到日本,但目前日本的盆景艺术闻名于世界,“我们却起码落后了20年”。而日本盆景之所以享有世界性的声誉,是以其原本特有的五针松、大阪松等优良树种为基础,进行市场化、产业化运作的结果。《盆景发展应从“小”做起”》是盆景业较为广泛的共识。但盆景是一种特殊的产品,选择优良而适宜的树种才是盆景小型化的关键因素。
江苏如皋好雨园景自1989年起开始对该物种的拯救、培育和利用工作。经过长达14a从高山至山脚再到平原等三个阶段的引种、驯化和繁育,至2003年,通过无性扩繁,使植株总量达到100余万株,建立了国内唯一大规模的珍珠黄杨人工种群,并初步选育过五个优良无性系,为新品种的选育提供了丰富的材料基础。好雨黄杨,乃是2003年在此种群中发现的、具有显著差异的无性系品种。其后,经过17a的对比观察,和最近四年共4批次的繁育和抗逆性栽培试验,结果证实,好雨黄杨与珍珠黄杨其他品种相比,具有明显的特异性、高度的一致性和稳定性。
好雨黄杨的培育成功,不仅对物种保护领域是一个贡献,更为满足国际国内盆景市场小型化的需求,提供了一个无与伦比的新优品种。

珍珠黄杨原名“凌霄树”。始初发现于安徽天柱山,以为“天柱山所独有植物”,人称天柱奇木。因在高山峭壁之上仰天破云而生,故而得名。作为典型的高山常绿树种, 珍珠黄杨性喜温暖湿润气候及酸性土壤。木质细腻坚实、叶片革质光亮,水份消耗少。耐旱,耐荫,抗严寒,生长极慢是其最为显著的特点。
多年的调研与实践经验表明,野生珍珠黄杨下山后难以成活的主要原因,是其特殊的生存环境发生突然变化所至。与其他生命体一样,野生珍珠黄杨在其漫长的生态历史过程中形成了自己特有的生长惯性。即在特定自然环境中,习惯按照自身的需求和方式,来调节细胞组织的适应功能,吸收外部养分,以维持其生命成长。改变了这种惯性,就极易导致生长不良甚至死亡。道理跟我们人类常见的“高原反应”、“水土不服”几乎没什么两样。
野生珍珠黄杨对环境要求极为严格:要求土壤营养丰富、通透良好、湿润并呈微酸性,空气湿度大,日光温和,忌夏日暴晒。一般来说,下山珍珠黄杨根据树龄和根系受损程度的不等,在移栽后的数月甚至一两年内,只要能给予适当的水份并精心护理,表面看来枝叶并不会发生太大的变化。树龄越小,根系细胞的的活力较强,维持生长的时间则稍长。但即使暂时得以存活,往往表现为春季返青迟缓,休眠期长,枝叶稀疏薄弱,多处于僵而不死的状态。树龄越大,则根系萎缩越快,尽管主干中的养分仍可维持叶片不致立刻枯萎,甚至偶有新根发生,但生长态势仍会随着时间的推移,逐渐衰减。
根据以往的资料及此次的调查结果,驯化成功的如皋珍珠黄杨原有无性系品种在适应性及抗逆性上与野生种相比有较大提高。而好雨珍珠黄杨,是在以大别山珍珠黄杨为原始材料,通过近20年连续不断继代扦插驯化,和培育出大规模人工种群的基础上,发现和培育成功的珍珠黄杨新品种。 在区域上经历了由高山至山脚再到平原的过程,在栽培方法上经历了由精细到粗放的阶段,并完全采用露天自然条件下的栽培模式,经受过冬季冰雪、夏季烈日以及雨涝干旱等恶劣气候的考验,具有较强的适应性。其树种特性、生长态势、抗逆性能以及栽培技术等与野生珍珠黄杨均有着明显的区别。
在地栽或盆栽条件下,新品种好雨黄杨具备以下生长特性: 1.耐阴喜光,在一般室内外条件下均可保持生长良好。但长期荫蔽环境中,叶片虽可保持翠绿,但生长态势会稍有减弱。 2.喜湿润,可耐连续一月左右的阴雨天气,但忌长时间积水。 3.耐旱,只要地表土壤或盆土不至完全干透,无异常表现。 4.耐热耐寒,可经受夏日暴晒和耐-20。C左右度的低温。 5.对土壤要求不严,以轻松肥沃的沙质壤土为佳,盆栽亦可以蛭石、泥炭或土壤配合使用,耐碱性较强。

人类文明的起源和发展总是和植物的驯化联系在一起,植物以人工栽培方式的出现是人类自身发展史上第一次伟大的革命。植物引种驯化(introduction and acclimatization)是一个庞大的理论与实践课题(廖馥荪,1966)。驯化则是人类对植物本性的主动改造过程,它可以在植物的系统发育中和个体发育中完成,植物最后产生了适应于新的自然环境条件下的新特性和新性状。
引种驯化的方法概括地可以分为两大类:简单引种:在相同气候带(如温带、亚热带、热带)内或差异不太大的条件下进行相互引种的方法,它不需要经过驯化阶段就可引种成功。复杂引种:在不同气候带之间以及气候差异较大的地区之间进行相互引种的方法。复杂引种难度较大,要通过改造性的栽培技术措施才能最终成功。通常采用下面几种技术路线进行:1)处理种苗;2)逐渐引种;3)驯化幼苗;4)继代扦插;5)多代连续驯化;6)选择育种。珍珠黄杨是一种分布于中国亚热带高山悬崖地区的珍稀濒危物种,它自然分布在海拔1200-3000 m悬崖生境,对生境要求比较严格,喜欢温暖湿润气候(郑勉和闵天禄,1980)。很多被盗采的珍珠黄杨下山后的命运,显明的证实了简单引种是无法达到理想的栽培效果的。
江苏如皋好雨园景自1989年起开始对珍珠黄杨的调研与拯救工作,经过长达30年从高山至山脚再到平原等三个阶段的引种、驯化与繁育,成功选优繁育出新一代新品种——好雨珍珠黄杨。好雨黄杨不仅从根本上摒弃了珍珠黄杨原品种生长 过于缓慢、枝条极易萎缩、盆栽难以存活等缺陷,而且保留和大幅提升了珍珠黄 杨原品种优良部分的特性,休眠期短,长势强盛。成为人类唯一真正可利用的珍珠黄杨种质资源。
后来有研究表明:同样一个无性系,经多代扦插,其扦插生根率有逐代提高的趋势,无性系继代苗的当年生侧枝,扦插成活率高于原株上的扦插成活率,而继一代与继二代无性系插穗,扦插苗成活率基本接近(周盛,1994)。另一些学者的研究证实插穗生根成活率与MIn(内源抑制物质)含量呈负相关倾向,并且MIn含量随母株年龄递增,而反复继代扦插可延缓母株老化,使母株复壮幼化,增强对环境的适应力(季孔庶,1999)。珍珠黄杨的驯化培育过程,是在一个逐步远离于高山气候,而在较为广域的人居环境条件下,通过继代扦插并选优汰劣,从而获得一个能够适宜人们栽培和观赏、性状稳定、抗性较强的优良种群。
参考文献:南京林业大学《珍珠黄杨遗传多样性及其资源保护及利用》黄焱 (2008)
翟锦如 2021.2
关键词:变异机理 分布区域 繁殖方法 表型差异 品种选育

珍珠黄杨的表型差异,是指珍珠黄杨的植株形态、叶片颜色和大小,以及枝节长度等质和量可视性差别。
一个物种的遗传结构受到其交配系统、基因流、种子散播机制、繁育方式和自然选择等一系列进化因子的影响,而繁育方式则是确定植物群体遗传结构最重要的因子(Hamrick & Godt,1989)。珍珠黄杨是典型的异花授粉植物,这就决定了其群体内多样性程度远大于群体间。此外,珍珠黄杨在长期的生长和演化过程中,对不同环境会形成不同的适应机制,从而产生从器官、个体到群体水平的各种变异现象。
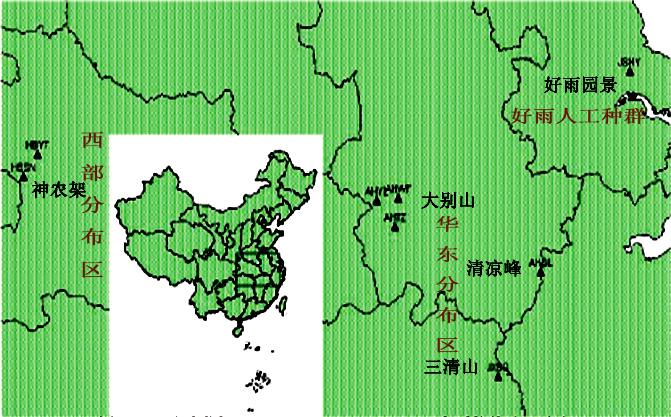
2008年,南京林业大学根据珍珠黄杨主要性状的差异,对珍珠黄杨分布的7个自然群体和一个人工栽培群体,选取17个有分类价值的枝叶性状进行观测分析。结果证实,珍珠黄杨随着分布区域不同,有着明显的表型差异。各群体中AHQL和HBSN这2个群体内形态变异程度要明显高于其它群体;而AHWF和 AHYL 的形态变异程度最低。基于珍珠黄杨8个群体17个表型性状的欧氏距离聚类分析结果表明:以阈值18为分界线,可将珍珠黄杨8个群体分为3类,AHTZ、HBYT、AHQL 和AHWF 为一类,AHYL和JSHY为一类,而 JXSQ和HBSN为另一类。【详细请参阅《珍珠黄杨遗传多样性及其资源保护及利用》黄焱 (2008))】

多年的试验及调研结果表明,由于珍珠黄杨遗传多样性丰富复杂,对环境较为敏感,通过种子繁殖的后代极易产生叶片变大,植株变高,枝条变长,树型松散等"逆向变异",从而失去其本有的形态特征和应有的观赏价值。而通过无性繁殖,虽然保留了珍珠黄杨的观赏特性,生长特性却难以改变。并且,除新近培育成功的“好雨”新品种以外,其他几乎所有珍珠黄杨类型,对环境要求均较高,尤其是生长过程中枝条萎缩现象,也很难改变。

珍珠黄杨表型变异的价值 珍珠黄杨为中国特有的濒危珍稀树种,主要分布在亚热带高山悬崖环境上,其分布区 地形复杂,气候和土壤条件千变万化,以及通过长期的地理隔离、自然选择和人工选择(栽 培群体),使其产生了极其丰富多样的种内变异。表型变异是遗传型和环境因子共同作用 的结果,表型变异蕴藏着遗传变异(Zhang,1998)。表型变异越大,可能存在的遗传变 异越大。研究自然存在的表型变异,是研究遗传变异的先导。
国家林业局新闻时空《南通市成功引种与驯化珍珠黄杨》2005.7.1 , 南京林业大学 《珍珠黄杨优良无性系形态特征分析》黄焱 翟锦如 季孔庶(2005)、《珍珠黄杨不同扦插继代的荧光特性》黄焱 季孔庶 汤庚国(2008),《珍珠黄杨遗传多样性及其资源保护及利用》黄焱 (2008)、《珍珠黄杨春季扦插生根性状差异及内源激素变化》黄焱 季孔庶 方彦 翟锦如(2007)、《珍珠黄杨开花及调控研究》周丹丹(2009),《珍珠黄杨叶片的蛋白质提取方法探讨》王明娟 季孔庶(2009),《珍珠黄杨 DNA 的提取方法比较及 ISSR 反应体系的优化》 吕乐燕 季孔庶 黄焱(2007), 《 珍珠黄杨组织培养及种子休眠解除技术的研究》牛琳(2007)





 苏ICP备05010171号
苏ICP备05010171号